
Though it is unfashionable these days to think of organisms as machines, to me Williams's fox is a matter-energy transfer machine that needs to catch and devour Williams's rabbit merely to continue to exist. Williams thinks that the fox eats the rabbit in order to pass on its genes. Dawkins would call Wiliams's fox a vehicle for its own genes' desires for replication. I, on the contrary, see reproduction as a physiological luxury rather than an imperative that is necessary for that fox to go on living. In my view, if and only if that fox's economic life is going well can it afford to reproduce. Is it fair to say that life is "all about" reproduction and the handing down of genes from one generation to the next?
(Niles Eldredge. Reinventing Darwin. Weidenfeld & Nicolson. 1995)
Idle Theory agrees, and goes further. Excessive reproduction - reproduction that increases population - is a threat to life. As their numbers rise they face the twin threat of overgrazing their habitat and becoming targets for predators. They would leave easier and safer lives if they restricted their reproduction to a minimum.
Asexual Reproduction
In Idle Theory, reproduction is regarded as a special case of growth. The creatures grow because they slightly overmaintain themselves, producing more replacement parts than are needed. Unicellular organisms then divide when they reach a critical size. Multicellulars made up of dividing unicellulars stop growing after reaching a maximum size, and the excess cell population is either lost, or is bundled together into a new baby multicellular which then begins an independent life.
This kind of reproduction is asexual. Only one parent is needed. Dividing cells carry the same DNA, and the offspring of asexually reproducing creatures (which include many plants, and several animals, including some types of lizards) are genetically identical to their mothers.
And this asexual reproduction is automatic. It just happens. When a growing cell reaches a critical size, it divides into two. In Idle Theory, cells reproduce like drops of water from a leaking tap. The creatures have no wish to reproduce: they just do, helplessly. The real problem for life is how to screw down the leaking reproductive tap.
Multicellular creatures grow by mitotic division from a single cell. If cell division just keeps happening, a multicellular will just get bigger and bigger. If there is an optimum size at which multicellular idleness reaches a maximum, and any bigger or smaller means lower idleness, then unrestricted cell division will rapidly increase its size beyond the optimun, into life-threatening low idleness.
Mitotic cell division produces two genetically identical cells, which then go on to divide again. But another form of cell division, meiosis, produces four cells, each of which contains half the number of chromosomes of a normal cell. These sex cells only develop and begin to divide when the full complement of chromosomes is produced by uniting with another sex cell.
The effect of meiosis, in the absence of union with other sex cells, is to produce non-dividing cells. Meiosis, therefore, is the end of the line for cell division. Meiosis stops further cell division - so long as haploid sex cells are prevented from uniting, and kept separate and apart. If a mitotically-dividing multicellular which is grow at 1% per day can induce 1% of its cells to divide meiotically rather than mitotically, it can stop further growth. A multicellular can then grow up to its optimum size, and then switch some cell division from mitosis to meiosis.
In the body of the multicellular creature, mitotic division would continue, with just enough new cells replacing dying cells. A mitotically dividing cell would leave many descendants: a meiotically dividing cell would leave none.
Sexual Reproduction
Sexual reproduction would then only occur if sex cells could manage somehow to unite. If the excess population of cells was simply discharged from the multicellular, a few sex cells could fuse with other sex cells and begin building a new multicellular. Once this kind of process is set going, reproduction becomes sexual. Multicellulars grow to an optimum size, and then begin discharging haploid non-dividing cells, a very small number of which unite with other haploid sex cells, and begin dividing and growing to build another multicellular.
What is the value of sexual reproduction? One common explanation is that sexual reproduction, which takes half the genes from the mother, and half from the father, results in diversity. No two offspring are genetically identical, and the slight genetic differences make for different fitness. A sexually producing population has enough inbuilt variety inherent in it to be able to respond to changing circumstnces in ways that asexually reproducing populatioms cannot.
The vegetative progeny of an individual (i.e. progeny produced without meiosis and subsequent fertilization) are genetically identical with each other and with their parent, unless a mutation has occurred. Suppose that among the vegetative descendants of a single individual ten different mutations have occurred, at different loci and in different lines of descent. Then there will exist ten different genotypes upon which selection can act. If however, these ten lines could cross sexually, three different genotypes would be possible at each locus (AA,Aa,aa) and these could be combined in any manner, giving a total of 310 = 59,049 different possible genotypes.
(Maynard Smith. The Theory of Evolution. Ch.12)
In the orthodox view, reproduction is pretty much the primary purpose of the creatures, to the point that life itself is almost defined as reproduction. But, in Idle Theory, as for Niles Eldredge, reproduction is an expensive luxury. Accordingly, Idle Theory argues that, rather than try to reproduce as rapidly as possible, the creatures are generally trying to reproduce as slowly as possible. The principal exception to this rule occurs when a population suffers from predator attack, and responds by increasing its reproduction rate.
If the creatures are mostly trying to slow up their reproduction rate, then a different quite explanation of sexual reproduction can be offered: that sexual reproduction, by making reproduction more complicated and difficult, - particularly if lengthy courtship rituals are also required - acts to impede reproduction. Sex is itself a contraceptive device, devised to limit automatic asexual reproduction. Any number of hurdles have to be overcome for sexual reproduction to work.
- Meiotic division of cells into non-dividing sex cells ensures that most sex cells (eggs and sperm) never reproduce. If sex cells are to divide, haploid female sex cells must fuse with haploid male sex cells to produce a diploid mitotically dividing zygote.
- Female sex cells are kept separate from male sex cells. Females have seek out males to fertilise their eggs. Males and females may not encounter each other.
- When males meet females, they may have to go through elaborate and lengthy courtship rituals, which may be unsuccessful.
- When males have met females, they have to successfully mate. Male sperm cells must perform navigational feats to reach the female egg.
- The fertilized egg, or zygote, has to develop successfully to term. A novel combination of genes, or environmental conditions, may thwart this.
- The independent offspring has to survive to maturity before, as an adult, it begins to produce sex cells, and can engage in sexual reproduction.
Thus, if asexually reproducing creatures can be taken to regularly divide or produce offspring, sexual reproducers face a whole string of obstacles.
Sex Ratio Reproduction Control
In The Origin of Species, Darwin gives a famous example of the effect of even slow reproduction:
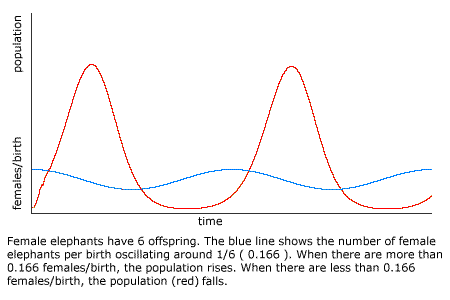
The elephant is reckoned to be the slowest breeder of all known animals, and I have taken some pains to estimate its probable minimum rate of natural increase: it will be under the mark to assume that it breeds when thirty years old, and goes on breeding until ninety years old, bringing forth three pair of young in this interval; if this be so, at the end of the fifth century there would be alive fifteen million elephants, descended from the first pair.
(Darwin. On the Origin of Species. Ist Ed. Ch 3.)
Darwin's elephants
But in a sexually reproducing population, there could be any number of elephants alive after five centuries. If the first pair give birth to six daughters or six sons, and elephants die at the age of one hundred years, then there will be no elephants at all. If female elephants always produce five sons and one daughter, then there may be no more elephants after five centuries than there were at the outset (neglecting any inbreeding problems). Only if female elephants each produce two or more daughters will there be an expansion in the population of elephants.
Sexual reproduction also offers the possibility not just of restricting reproduction, but of exercising a high degree of control over reproduction, both increasing or decreasing it. This can be achieved if the ratio of males to females can be varied.
If a reproducing population consists mostly of females, with a small number of males, then the few males can fertilise all the females, the population can double in whatever period it takes for the females to bring offspring to term.
If a reproducing population consists mostly of males, with a small number of females, then the population may grow slowly, or not at all. In fact, the population may actually dwindle, as the few reproducing females are unable to maintain numbers.
This facility to control population could be extremely useful. If, under predation, a population shifted towards producing more females than males, then it would be more able to counteract losses from predation. But when, without predators, a population threatened to overgraze its environment, the production of more males than females would act to slow, or reverse, population growth.
The sex of crocodiles is determined by the depth at which eggs are laid. Thus crocodiles can, in principle at least, control the sex ratio of their populations,
Effective Sex Ratios
Since part of the process of sexual reproduction requires the active participation of the creatures themselves - meeting and mating -, the creatures are able to exercise control over reproduction.
Where a sexually reproducing population has no control over its actual sex ratio, it can still control its population if some members abstain from sex, and become non-reproducers. Then, whatever the actual sex ratio, the effective sex ratio is determined by the proportions of each sex which do not abstain from sex.
The rate at which the population grows depends on the number of sexually active females. If all males mate with a single sexually active female, then population will increase at the rate that the single female gives birth. If all females mate with a single sexually active male, the population grows at the rate each female gives birth times the number of females. It is the sexually active female population that determines population behaviour.
Since the population is determined by the number of active females, it is female sexual behaviour which really matters most. In a population of 10 females and 10 males, then if 9 females abstain from sex, only one female remains effectively fertile, and there are 1/10th of the number of births there would otherwise be. But if 9 males abstain from sex, unless there are strict monogamy rules, the single remaining sexually active male will quite likely be able to fertilize all 10 females, and maximum population growth results. Female abstention has effects on the population in proportion to the number of abstaining females, but male abstention can only have an effect on population if all males without exception abstain.
If selection pressure on a sexually reproducing population is for minimum reproduction, then changes in female behaviour have much more effect than changes in male behaviour. So the likelihood is that if abstention is practised, it will be practised by females. It will tend to be females, not males, who will be disinclined to mate. If males are disinclined to mate, while females are inclined, it only takes a small minority of rogue males to keep reproduction levels high.
If females are generally disinclined to mate, while males are not, then females will determine which males, if any, they will mate with. Selection will favour those males which are able to overcome female disinclination. It may be that it is this kind of selection pressure which produces glamorous peacocks and drab peahens, or extraordinary male birds of paradise.
Under predation, or some other environmental hazard, selection favours increased reproduction. This translates into a high proportion of females to males, or a larger number of sexually active females. Males, spoilt for choice, become drab, and females become glamorous.
Conclusion
Sexual reproduction not only allows useful genes to be rapidly dispersed through a population, but it also allows control to be exercised over population levels, either by a change in the actual ratio of the sexes, or by an effective change in the sex ratio through the sexual inactivity of otherwise fertile members of the population. Sexually reproducing populations can act very rapidly to either expand their numbers or reduce them.
Biologists for whom reproduction is the principal imperative of life - "breed early, and breed often" - only see one side of the picture. In some circumstances, it is quite true that populations subject to predation, or disease, or some other hazard, must increase their reproduction rates if they are to survive. But it is equally true that where a population is not subject to predation, disease, and similar hazards, then the danger it faces is one of overpopulation, and selection pressure must be to reduce reproduction.