Multicellular life:
Once these primitive cells have begun dividing and multiplying in streams and pools of nutrients, they may simply disperse and continue to live as autonomous entities. But it is possible that rather than a dispersing cloud of autonomous cells, there could appear aggregations of cells, much as cells are here regarded as aggregations of primitives. Two kinds of multicellular organization are considered. In the first, coercive type, cells feed off each other, parasitically, and the idleness of some cells is increased at the expense of others. In the second, cooperative type, cells act in concert in ways which increase their idleness.
1. Coercive multicellular life - Parasites and Predators:
To each primitive cell, the food available would appear to consist of both the nutrient pool and the cell population. To a cell, other cells which store nutrients would appear as local concentrations of energy. Thus cells could become predators or parasites upon other cells. Where two cells are adjacent to each other, one cell may be able to extract nutrients from the other. If the nutrient concentration in the cell is higher than in the cell external environment, a cell that extracted energy from other cells would be able to operate at a higher idleness than otherwise, assuming that cells had no defences.
Where one cell becomes parasitic upon another, the extractor idleness of the parasite increases, as it does less work feeding from the energy-rich host than it would drawing sustenance from the less rich environment. At the same time, the extractor idleness of the host cell falls, because the extractors now have to supply enough energy to power two cells rather than just one.
If, for simplicity, it is assumed that this extra burden consists of Pm + Pr for each parasite, then host extractor idleness becomes
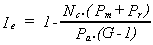
where Nc is the number of cells in the chain. Where there is one parasite cell, Nc = 2. If host extractor idleness remains greater than zero, the host survives. If a parasite reduces host idleness to zero, the host is unable to maintain itself, and disintegrates or dies. In this latter case, the parasite is effectively a predator that kills its prey.
 If host extractor idleness is high, it may be able to support a chain
of parasites. This may arise if the parasite itself becomes parasitizes
by a third cell, and so on.
If host extractor idleness is high, it may be able to support a chain
of parasites. This may arise if the parasite itself becomes parasitizes
by a third cell, and so on.
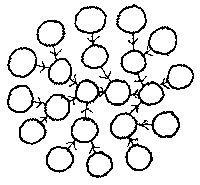 There can be any number of configurations of host cells which
extract energy from the environment, and parasite cells which extract
energy from host cells. They could, for example, form spheres whose
surface cells fed extracted energy from the environment, and whose
interior cells were chains of parasitic cells taking energy from surface
cells.
There can be any number of configurations of host cells which
extract energy from the environment, and parasite cells which extract
energy from host cells. They could, for example, form spheres whose
surface cells fed extracted energy from the environment, and whose
interior cells were chains of parasitic cells taking energy from surface
cells.
In the case where parasitic cells form a growing chain or sphere, the idleness of the host cells falls, and growth stops when terminal or surface cell idleness approaches zero. A single host can only support so many parasites. When host cells reach zero idleness, and disintegrate, either the entire dependent chain of parasites disintegrates, and all cells in it die, or else the terminal dead cell is shed, and the next cell in the chain either reverts to feeding from the environment or else latches onto another cell. This cell may itself be part of a parasitic chain. This chain would then find its stored energy being rapidly sucked out by the predator, and disintegrate.
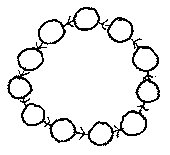 If the cell happens to latch onto the tail of the chain of
parasitic cells dependent upon it, biting its own tail, then the energy
stored in the chain dwindles away rapidly, and all cells in the chain die.
Within the body of a system of parasitic cells, therefore, there would
be a tendency for circular channels to form, where parasitic cell chains
had wrapped around to feed on themselves, died, and disintegrated,
leaving these channels.
If the cell happens to latch onto the tail of the chain of
parasitic cells dependent upon it, biting its own tail, then the energy
stored in the chain dwindles away rapidly, and all cells in the chain die.
Within the body of a system of parasitic cells, therefore, there would
be a tendency for circular channels to form, where parasitic cell chains
had wrapped around to feed on themselves, died, and disintegrated,
leaving these channels.
The problem for such parasitic systems is that while the parasites enjoy increased idleness, the idleness of their hosts is diminished, and the real or effective idleness of such systems is that of their least idle member. In parasitic systems, this lowest idleness must always be less than the idleness of free-living, autonomous cells. All that happens in parasitic systems is that the idleness of some cells is increased at the expense of others. There is no net increase in idleness.
In high energy environments, where single cells all have high idleness, predation and parasitism is unlikely to occur, because there may be effectively no gain in idleness from adopting a parasitic or predatory mode of existence. However, where environmental energy density falls, and some cell types are driven down towards zero idleness, parasitism and predation may offer the only viable mode of existence. Predators and parasites are liable to emerge among the least idle types of cell, with the most idle cells as their prey or hosts.
Cooperative multicellular life:
If the pool or stream environment in which the cells multiply is in some degree corrosive, so that cells are having to perform a great deal of work replacing components in their external extractor shell, then if cells can bunch together, and share "party walls" rather like semi-detached houses or terraces, then party walls which are not exposed to the corrosive external environment will require less maintenance. Maintenance energy costs will fall, and both assembler and extractor idleness will increase. In this case, each cell extracts energy from the environment, rather than adjacent cells, to power its maintenance processes. Thus each cell must have access to the external environment, and have at least one external wall. If a cell gets buried under other cells, losing access to the external environment, it must become a predator or parasite on adjacent cells.
There are many possible arrangements for such cells. If, for the purposes of this discussion, cells are assumed to have a cubic shape, which allows close packing. Various different configurations give different numbers of external faces, from 6 with a single cell to 1 with a double layer sheet of cells.
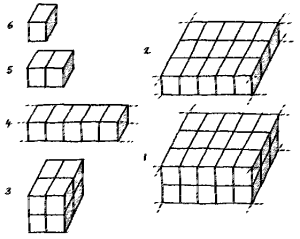 |
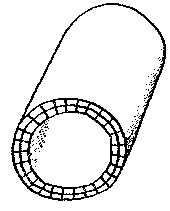 |
The optimum system, a bilayer sheet, has edges where each cell has 2 external faces, and corners with 3 external faces. If the sheet wraps round to produce a tube, all the corners, and half the edges, disappear, leaving two edges where cells have 2 external faces.
This tubular arrangement has fewest external faces if the tube is long and narrow, rather than short and wide. However, within a long narrow tube, the energy density of the medium is likely to fall, unless there is a flow of nutrients through the tube. If energy density within the tube is allowed to fall, cells inside the tube near the centre are liable to become parasitic on their more idle neighbours on the exterior of the tube. If this results in the death of cells at the centre of the tube, the two halves of the tube will separate; the tube will divide.
One possible solution to the problem of maintaining a flow of nutrients to the cells lining the interior of the tube would be for the cells along the tube to sequentially expand and contract - if they possess this capability -, and generate a kind of peristalsis, pumping the medium through the tube, and even generating forward motion for the entire tube, allowing it to move in the environment.
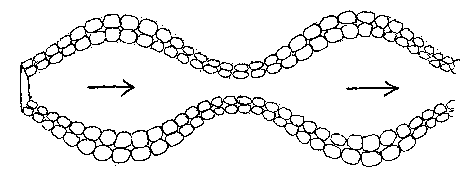
This would perhaps tend to provide a higher energy density inside the tube than on its external surface, as it sucked in not only nutrients in the environment, but single cells floating in that environment.
Once, for whatever reason, cells have formed into multicellular entities, other kinds of interaction become possible. Biological cells require a variety of nutrients - oxygen, water, sugars, amino acids, sunlight, carbon dioxide, trace elements -, and in a multicellular made up of some number of cells, some cells may have easier access to some of these than others. One side of a cell bi-layer may be able to easily extract nutrient A (e.g. oxygen) from the external enviroment, but not nutrient B (e.g. water) On the other side, the reverse might be the case. In this circumstance, it may prove easier - i.e. result in higher idleness - for one side to acquire nutrient B from its B-rich neighbour, and for the other side to get nutrient A from its A-rich neighbour. This could be regarded as mutual parasitism, or as a form of trade.
With this arrangement, cells that find it easy to extract nutrient A from the environment become the suppliers of A to the entire community, and cells that can easily extract nutrient B become general purveyors of B. At the same time, those cells which find difficulty in acquiring A or B from the environment entirely stop doing so, and take all they require from their neighbours. In this way, cells become specialized.
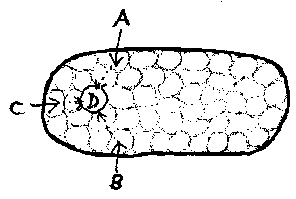
A similar argument can be applied to the production by cells of components or products which are required by all cells. If cells require A, B, and C in order to synthesize a product D, then assuming that A, B, and C are materials extracted from the environment by specialized cells at different locations in the multicellular, and are passed from cell to cell, then the optimum site or region for synthesizing D will be where the energy costs of transport are at a minimum. This sites, or region, could then become a specialized producer of product D, which is then transported out to other cells, which stop producing it. If there is no location where D can be produced with least energy, no specialization occurs, and all cells make it themselves.
The same kind of logic could apply where a multicellular is subjected to compressive or tensile forces. Where compression is at a maximum, cells may be assumed to counter this by constructing compression-bearing internal structures (e.g. bone) depending on the degree of compression. In doing so, they may become compression-bearing specialist cells (e.g. bone cells) which perform compression-bearing work not only for themselves, but the entire community of cells.
Once particular cells or groups of cells become specialized producers, economies of scale become possible. If, for example, a particular production process is most energy-efficient at a particular temperature, then N dispersed single cell producers, each losing heat from 6 faces ( a cubic form with unit side length is assumed ) will have a total heat loss, Q1, given by

where T1 is the cell temperature, and T2 is the temperature of the cell environment, and k is a thermal conductivity coefficient. Where the same N cells form a single cube, the side length of this cube is N1/3, and the heat loss Q2 is given by
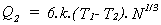
Thus the energy consumption of a 1000-cell cube will be 1/100th that of 1000 dispersed cells in the same environment, and each cell will require less energy to produce the same product.
If such economies of scale can be found to apply for various other activities - pumping fluids in tubes (blood vessels), oxygen induction (lungs), mechanical transport (legs) - then increases in size bring greater energy efficiency, and increased idleness.
What began as a collection of a few cells here first becomes a tube, and some cells along the length of the tube become specialized muscle cells which act to force ingested material along the gut tube, where other cells extract and distribute nutrients to other cells. Cells at the surface become specialized in providing protection against a corrosive environment. Cells between skin surface and gut specialize in producing enzymes which aid digestion, or act as energy stores. Each cell has its own particular task to perform. Each capability of a generalized all-purpose cell becomes the specialist activity of groups of cells - organs -, with location being the principle determinant of the specialisation adopted by a cell.

At the same time that hundreds or thousands of cells begin to form a multicellular entity which can move in the environment, feeding at different locations, it becomes necessary to coordinate activities. A great many activities may simply be under local control. But where a complex multicellular is to move around an environment, a large number of muscle cells have to act in concert, and some sort of control system must direct the entity in its path, if it is not to simply go in circles.
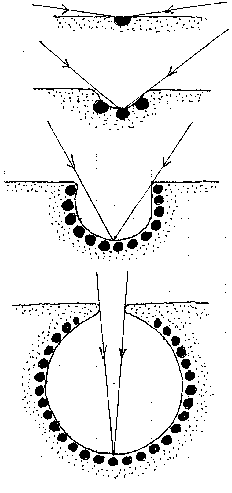 If one of the capabilities of a cell is to be respond to light (as plant cells can) or darkness by producing or not producing some product which is distributed throughout the multicellular causing muscle cells to contract or relax, then changes in external environmental light levels can, in principle, affect the behaviour of the entity. With two light-responsive cells on each side of the front end of the entity, when light falls on one, but not the other, the first produces the muscle-contracting agent, and the other does not. The result is that the entity turns towards the light.
Sensitivity to light levels can then be enhanced by increasing the number of light-responsive cells. Some cells may be responsive to light of different colours. Such a simple form of vision would only report overall light levels, and not a detailed picture of the external environment.
But if the light-responsive cells are sunk into a pit with a narrow exterior opening, so that they respond to light coming from a particular direction, then the field of vision of each cell becomes mapped onto the external environment.
In the natural world, two kinds of eyes have evolved - the concave and the convex. The concave eye, found in most mammals and reptiles and birds, is a single cavity with a single opening to the external world, whose interior is lined with light-responsive cells. Each of these cells responds only to light coming from a particular direction.
The evolution of this eye can be regarded as a gradual increase in resolving power. The first eye, a single light-responsive cell, can only distinguish between light and dark. As the number of cells in the eye increases, and the neck of the cavity narrows, cells respond only to events within a narrower field of view. At this point, large objects can be distinguished, because they appear to some cells, but not to others. However, increases in resolution are paid for with decreased light intensity as the eye's aperture decreases. The way round this is to introduce a lens which allows the aperture to be widened, and more light gathered. Then resolving power is determined by how closely light-sensitive cells can be packed on the retina.
The need for increased resolving power can be seen as a result of falling nutrient densities. If nutrients are visible objects, then when there is a high concentration, and the objects are nearby, a low resolution eye is quite adequate to discern them. When nutrients are widely dispersed, far away objects, a higher resolution eye is needed. The same applies for other sensors, such as an ear or a nose. In an extremely energy-rich environment, a primordial soup, living entities would have no need of such acute sensors: they would -- as it were - just open their mouths and swallow. It is only when the environment becomes energy-poor, when nutrients are widely dispersed, that such sensory acuity becomes indispensable.
The more acute the sensors becomes, the more detailed the information it provides, the more sophisticated the control system must be to 'makes sense' of what is seen, and translate this into an appropriate response. For a simple entity with a single cell eye, and a single cell nose, the conjunction of 'green' and 'sweet' may be sufficient to trigger eating the sweet, green apple directly in front of it. But when increasing acuity of vision results in seeing several apples, or several fuzzy green-applish blobs, at the same time, it becomes important to decide which way to go.
Increasing visual acuity also allows the possibility of constructing maps of the environment, which can be remembered. Rather than rediscovering the world every day, an animal with a memory knows where it is.
For these purposes, some cells have to combine with others in a central nervous system, which receives information from sensors - eyes, nose, ears - and activates muscles in response. And rather as light-sensitive cells can combine together to form an eye which can see far more clearly and acutely than a single cell, so also unspecialized cells that possess rudimentary abilities to store materials, and to switch processes on and off, become specialized as switches and stores in logic circuits made up of hundreds, even millions of cells, and whose combination provides computing and memory capabilities far in excess of any single cell. This brain is able to construct an image of the outer world, and to activate and de-activate a large number of muscles and other processes, using a variety of strategies.
In the simpler forms of life these strategies - which have the form If X Happens, Then Do Y - might well be fixed. It always does the same thing, given the same circumstances, its strategies as hard wired as the light receptors in its eyes. Move towards the first green thing you see, and eat it.
But if it uses soft strategies, which can be modified or replaced, then it has the possibility of learning new strategies. Rather than heading for the first green plant sighted, it may instead move towards the nearest plant, or towards the region where plants are most abundant, or where a particular type of plant is plentiful. New strategies can be proposed and tested, in a process of trial and error. If a strategy increases idleness, then it is accepted. If it decreases idleness, it is rejected. The converse value system - to act so as to minimize idleness - would ensure extinction.
The ability to propose a variety of different strategies, different courses of action, implies an ability to choose between them, to value one course of action above another. In an energy-rich environment, where food is abundant, one strategy is likely to be as effective as another, and indiscriminate action is possible. But when food is scarce, choosing the best strategy - the course of action which maximizes idleness - becomes a matter of survival.
The ability to simply remember where food is likely to be found is itself a remarkable time-saver. An animal that has no memory must spend long periods searching for food, while one which remembers can go straight to that food. And one that remembers, and has a (remembered) map of the locality, can work out the shortest path between these remembered food stores.
This requires a further ability. Not simply to conceive of various different options, but to simulate performing them, rather than actually performing them, and selecting the most successful simulated activity. Trial and error is first conducted in imagination, before being tried in practice. Here physical work begins to be replaced by mental work: instead of actively performing proposed courses of action, the actions are performed in imagination, before one is selected and acted upon. A lot of sitting and thinking is done. In terms of physics, there is no difference between brain work and any other kind of work: an active brain is using energy just like any muscle: it is simply that the work being done is one juggling with ideas rather than juggling with physical objects.
In this approach, mind and brain have the same relation as computer software does to hardware. The hardware does not change, but is able to run different programs much as tape recorders can play different pieces of music. And this mind-brain is simply one organ in a cooperative system of organs, all of which are indispensable.
If one of the capabilities of a cell is to be respond to light (as plant cells can) or darkness by producing or not producing some product which is distributed throughout the multicellular causing muscle cells to contract or relax, then changes in external environmental light levels can, in principle, affect the behaviour of the entity. With two light-responsive cells on each side of the front end of the entity, when light falls on one, but not the other, the first produces the muscle-contracting agent, and the other does not. The result is that the entity turns towards the light.
Sensitivity to light levels can then be enhanced by increasing the number of light-responsive cells. Some cells may be responsive to light of different colours. Such a simple form of vision would only report overall light levels, and not a detailed picture of the external environment.
But if the light-responsive cells are sunk into a pit with a narrow exterior opening, so that they respond to light coming from a particular direction, then the field of vision of each cell becomes mapped onto the external environment.
In the natural world, two kinds of eyes have evolved - the concave and the convex. The concave eye, found in most mammals and reptiles and birds, is a single cavity with a single opening to the external world, whose interior is lined with light-responsive cells. Each of these cells responds only to light coming from a particular direction.
The evolution of this eye can be regarded as a gradual increase in resolving power. The first eye, a single light-responsive cell, can only distinguish between light and dark. As the number of cells in the eye increases, and the neck of the cavity narrows, cells respond only to events within a narrower field of view. At this point, large objects can be distinguished, because they appear to some cells, but not to others. However, increases in resolution are paid for with decreased light intensity as the eye's aperture decreases. The way round this is to introduce a lens which allows the aperture to be widened, and more light gathered. Then resolving power is determined by how closely light-sensitive cells can be packed on the retina.
The need for increased resolving power can be seen as a result of falling nutrient densities. If nutrients are visible objects, then when there is a high concentration, and the objects are nearby, a low resolution eye is quite adequate to discern them. When nutrients are widely dispersed, far away objects, a higher resolution eye is needed. The same applies for other sensors, such as an ear or a nose. In an extremely energy-rich environment, a primordial soup, living entities would have no need of such acute sensors: they would -- as it were - just open their mouths and swallow. It is only when the environment becomes energy-poor, when nutrients are widely dispersed, that such sensory acuity becomes indispensable.
The more acute the sensors becomes, the more detailed the information it provides, the more sophisticated the control system must be to 'makes sense' of what is seen, and translate this into an appropriate response. For a simple entity with a single cell eye, and a single cell nose, the conjunction of 'green' and 'sweet' may be sufficient to trigger eating the sweet, green apple directly in front of it. But when increasing acuity of vision results in seeing several apples, or several fuzzy green-applish blobs, at the same time, it becomes important to decide which way to go.
Increasing visual acuity also allows the possibility of constructing maps of the environment, which can be remembered. Rather than rediscovering the world every day, an animal with a memory knows where it is.
For these purposes, some cells have to combine with others in a central nervous system, which receives information from sensors - eyes, nose, ears - and activates muscles in response. And rather as light-sensitive cells can combine together to form an eye which can see far more clearly and acutely than a single cell, so also unspecialized cells that possess rudimentary abilities to store materials, and to switch processes on and off, become specialized as switches and stores in logic circuits made up of hundreds, even millions of cells, and whose combination provides computing and memory capabilities far in excess of any single cell. This brain is able to construct an image of the outer world, and to activate and de-activate a large number of muscles and other processes, using a variety of strategies.
In the simpler forms of life these strategies - which have the form If X Happens, Then Do Y - might well be fixed. It always does the same thing, given the same circumstances, its strategies as hard wired as the light receptors in its eyes. Move towards the first green thing you see, and eat it.
But if it uses soft strategies, which can be modified or replaced, then it has the possibility of learning new strategies. Rather than heading for the first green plant sighted, it may instead move towards the nearest plant, or towards the region where plants are most abundant, or where a particular type of plant is plentiful. New strategies can be proposed and tested, in a process of trial and error. If a strategy increases idleness, then it is accepted. If it decreases idleness, it is rejected. The converse value system - to act so as to minimize idleness - would ensure extinction.
The ability to propose a variety of different strategies, different courses of action, implies an ability to choose between them, to value one course of action above another. In an energy-rich environment, where food is abundant, one strategy is likely to be as effective as another, and indiscriminate action is possible. But when food is scarce, choosing the best strategy - the course of action which maximizes idleness - becomes a matter of survival.
The ability to simply remember where food is likely to be found is itself a remarkable time-saver. An animal that has no memory must spend long periods searching for food, while one which remembers can go straight to that food. And one that remembers, and has a (remembered) map of the locality, can work out the shortest path between these remembered food stores.
This requires a further ability. Not simply to conceive of various different options, but to simulate performing them, rather than actually performing them, and selecting the most successful simulated activity. Trial and error is first conducted in imagination, before being tried in practice. Here physical work begins to be replaced by mental work: instead of actively performing proposed courses of action, the actions are performed in imagination, before one is selected and acted upon. A lot of sitting and thinking is done. In terms of physics, there is no difference between brain work and any other kind of work: an active brain is using energy just like any muscle: it is simply that the work being done is one juggling with ideas rather than juggling with physical objects.
In this approach, mind and brain have the same relation as computer software does to hardware. The hardware does not change, but is able to run different programs much as tape recorders can play different pieces of music. And this mind-brain is simply one organ in a cooperative system of organs, all of which are indispensable.
Multicellular reproduction:
While single cells can reproduce simply by dividing in two, multicellulars may in many cases have too complex a geometry to allow this kind of reproduction. It is possible that multicellulars could be formed from a population of cells uniting into a single entity. But most biological multicellulars start life as a single cell which repeatedly divides and grows into the adult form, in which each cell gradually becomes a specialist in some necessary activity. If multicellulars can only be grown in this way, then there is a problem. This is that it has been argued that multicellulars are more idle than unicellulars, and that life tends to be difficult for unicellulars. Now, since a multicellular must start life as a single cell, this cell may quite simply be unable to survive in the environment. It is only when they have reached some threshold size that they become able to survive. The biological solution to this problem has been to create an artificial energy-rich environment in which the initial cell - the zygote - can survive and reproduce, and grow into a viable multicellular. In the eggs of birds and reptiles, or the seeds of plants, the zygote is provided with a large supply of nutrients encased in a shell. Within this artificial environment, the growing multicellular is able to develop some way towards its full adult size, so that when nutrients within the egg shell are exhausted, it is large enough to survive in the natural environment. Mammals do it a different way: the zygote grows within the energy-rich body of the adult multicellular, until it achieves a size where it can survive in the external environment. In both cases, the new multicellular only lives through the early stages of its life thanks to a bequest from its parent multicellular. This assistance may extend to protection and sustenance after the young multicellular has entered the external environment. This form of reproduction does not have to be sexual reproduction, where the genes of the offspring are taken partly from one parent, partly the other. In asexual reproduction, any non-specialized cell could be the zygote. The advantage of sexual reproduction is that each offspring is not identical to its parents, so that sexual reproduction results in a diverse variety of multicellulars, some of which are more idle, and therefore survive longer, than others. A multicellular with an intestinal tract and a blood stream carrying nutrients to every cell can be regarded as a population of cells multiplying in a stream of nutrients. The result is that this population reaches a maximum which is dependent on the power input to the stream. This climax population level corresponds to the adult size of the multicellular.
Aging as succession:
In the plant succession model, less idle fast reproducing plants are gradually replaced by more idle slow reproducing plants, until at climax only the slowest reproducers of all remain. This model can also be applied to the community of cells that make up a multicellular entity, which each draw nutrients from a circulating blood stream. A multicellular starts life as a single cell, which grows and divides into two daughter cells, which in turn grow and divide again. If it is assumed that some of the daughter cells divide grow and divide more rapidly, and some more slowly, than the first cell, the growing multicellular comes to be composed of a number of 'species' of cell, some reproducing rapidly, some very slowly. Initially, the cell population grows exponentially, and fast multiplying cells quickly predominate. Exponential growth ends when the population of cells reaches the limit determined by the energy flow in the blood stream. Given that the multicellular is composed of almost entirely of fast dividing cells, the limit population is low, and the multicellular is consequently small in size. Accidental damage entailing the death of numbers of cells is quickly repaired by replacement new cells. But since more idle slow reproducing cells can survive where fast reproducing cells cannot, they gradually come to predominate, and the limit population increases, so the multicellular continues to grow in size, but not exponentially. Mature adulthood corresponds to a climax population of very slow reproducing or even non-reproducing cells. This adult, however, is almost entirely unable to replace cells lost through accidental damage or disease. If the death rate of cells exceeds their birth rate, the cell population gradually falls, and the multicellular dwindles in size, wasting away. It is as if a climax forest of slow-growing trees becomes filled with clearings, where trees have toppled, and not been replaced because new trees grow so slowly. The dwindling number of remaining cells, furthermore, have to perform much the same amount of work as before: wasting bones have to carry the same weight, and wasting muscles perform the same work. The idleness of these remaining cells thus steadily decreases, and the multicellular dies when the cells of some vital organ reach zero idleness, and these cells die, and like falling dominoes, other dependent organs and cells also die in rapid succession.
The death of a multicellular is the death of its component cells. If the organ that fails is the heart, then the blood stream becomes a static nutrient pool. Depending on how much energy is stored in the pool, cells may live for some time, with extractor idleness falling as it becomes progressively more difficult to extract nutrients from the pool.
The converse process to succession can explain how aged parents manage to give birth to youthful offspring, rather than offspring as aged as themselves. The zygote may start off dividing very slowly within the artificial high-energy environment of egg or uterus. But if the growing population is made up of cells, some of which divide more rapidly, some more slowly, then the faster-dividing cells will rapidly predominate. The result is a fast-growing juvenile composed of mostly fast-reproducing cells. Once removed from the artificial high-energy environment, the juvenile begins to age, gradually becoming composed of slower and slower dividing cells.
Societies of multicellulars:
The process of evolution envisioned here is not one which aims at producing beautiful and elegant and high-minded forms of life, but one which is driven by steadily falling energy densities, ever scarcer food, and decreasing idleness, to adopt the ever more efficient organization. The step from single cell to multicellular cooperative increases idleness, as does the specialization of cells in an internal division of labour within a multicellular, while growth in size of organs and entire multicellulars brings economies of scale and otherwise increased performance.
Once cells have organized themselves into multicellular communities, and briefly increased their idleness, before the ever-worsening environmental circumstance erases this advantage, the next stage is the formation of cooperative societies of multicellulars, in a process that repeats the development of multicellular life from unicellular life. Individual multicellular forms of life become the component "cells" of a "multicellular" society. These societies, needless to say, do not exist for their own sake, but to serve the interests of their individual constituents, who in turn serve the interests of their constituent cells, and so on down.
It may be asked how it is, if there is such a constant need for life to organize itself first into complex multicellulars, and then into societies of multicellulars, that there are any unicellular forms of life still extant? Ought they not to be extinct? Several answers can be offered.
In the first place, even if unicellular organisms are taken to be inherently less idle than multicellulars, they have a great many more energy-rich nooks and crannies in which they can survive than their larger multicellular counterparts. In the second place, it may be that some unicellular forms of life are in fact so inherently idle that it has never been necessary for them to adopt a cooperative mode of existence, and it was only the least idle forms of unicellular life which were driven to a multicellular mode of existence under threat of extinction. There are some kinds of bacteria which derive their energy from the oxidization of iron - something few other unicellular organisms have managed to do -, and there is a lot of iron and oxygen in the universe. Equally, unicellular plants only require sunlight, water, and carbon dioxide, which are also abundant. Of the remainder, which require sugars and proteins as nutrients, a great many are parasitic upon larger multicellular forms of life. Every human being carries many different species of them, living off discarded skin, or within the intestinal tract. One of the most intensively studied - E. Coli - takes its name from the human colon in which it normally resides. Most of these parasites are harmless, living on human wastes rather than upon living human tissue. Others - bacterial infections - can multiply explosively in the energy-rich human internal environment, with fatal consequences for their hosts, and themselves.
There are many such natural societies. Many animals live in herds, or hunt in packs. Ants, termites, and bees have highly structured societies, with definite roles assigned to different members. From the point of view of Idle Theory, these do not appear as "advanced" forms of life, but rather as forms of life that have come so near to extinction that they have been forced into another level of integration which more successful - more idle - life forms have yet to encounter any need. For while ants and termites form societies, there are plenty of other insects - ranging from spiders to flies - which do not. And there are equally plenty of birds and mammals which live autonomous existences, able to fend for themselves.
The Long Run:
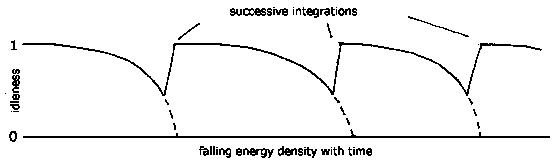
The overall vision of evolution proposed by Idle Theory is one of increasing complexity and integration. The first primitive lifeforms combine to form cells. and then the cells combine to form multicellular entities, and then the multi-cellulars combine to form societies or communities. Each step of integration results in increased idleness.
But all this takes place, and is ultimately driven by, a circumstance in which energy is becoming steadily more dispersed and diffuse in the environment. This is the operation of the Second Law of Thermodynamics, by which the entire universe cools, and becomes dispersed.
The most primitive forms of life, in the remote past, grew and multiplied in a high energy environment, at very high levels of idleness. As energy in the environment became more dispersed, their idleness fell. Only at this point did it become necessary for these primitive life forms to combine into more idle configurations. Those that could do this survived, while the remainder became extinct. In time, the further degradation of the environment required further integrations, to counteract the general trend of falling idleness. Thus the circumstance of life is that it is always becoming less idle, as energy becomes more dispersed, and every life form must work longer to maintain itself. Multiple successive integrations counteract this trend, and act to increase idleness. Life, therefore, always needs to come up with new devices and arrangements to increase idleness to reverse the general trend of decreasing idleness.